 |  |
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
BD Rhapsody

BD Rhapsody 技術(shù)利用卡式芯片在磁性的寡核苷酸條形碼標(biāo)記微球上實(shí)現(xiàn)單細(xì)胞捕獲和 mRNA 轉(zhuǎn)錄本的分子標(biāo)簽,然后將這些微球合并到單個(gè)管中用于 cDNA 擴(kuò)增和文庫(kù)構(gòu)建。可滿足 100~10000 個(gè)細(xì)胞的自動(dòng)分選、擴(kuò)增及建庫(kù)。
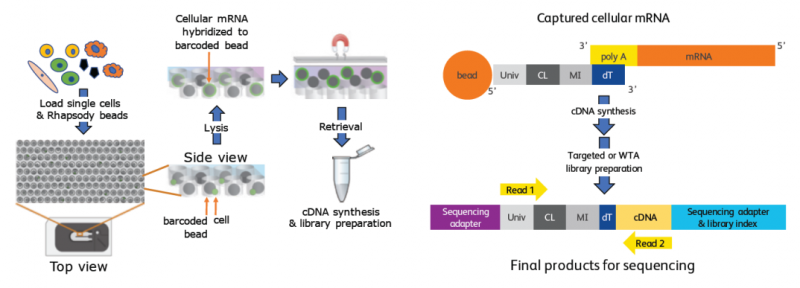
圖 BD Rhapsody 技術(shù)原理
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
此外,該技術(shù)使用帶有寡核苷酸的高質(zhì)量抗體(Ab-oligos),這條寡核苷酸帶有抗體特異的條形碼,細(xì)胞在經(jīng)過(guò) Ab-oligos 標(biāo)記后,可在單細(xì)胞水平同時(shí)獲得轉(zhuǎn)錄組和蛋白表達(dá)。
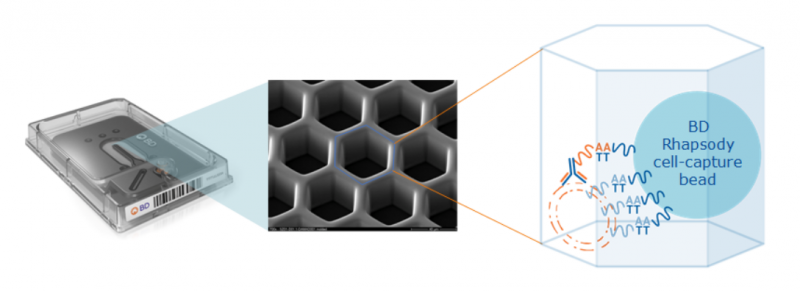
圖 單細(xì)胞 mRNA 與蛋白同時(shí)測(cè)序
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370

| 高 效 | 每個(gè)樣本可測(cè) 100~10000 個(gè)細(xì)胞; |
| 2 天內(nèi)完成細(xì)胞懸液制備、單細(xì)胞捕獲、擴(kuò)增以及建庫(kù)。 | |
| 靈 活 | 抗體標(biāo)簽技術(shù)可實(shí)現(xiàn)多樣本混合捕獲; |
| 可選擇全轉(zhuǎn)錄組測(cè)序或目標(biāo)基因測(cè)序。 | |
| 可靠 | 利用成像系統(tǒng)對(duì)單細(xì)胞捕獲過(guò)程進(jìn)行質(zhì)控; |
| 單細(xì)胞捕獲效率高達(dá) 80%,多細(xì)胞比例 <1%(1,000 個(gè)細(xì)胞中)。 | |
| 整合 | 可同時(shí)檢測(cè)單個(gè)細(xì)胞的 mRNA 水平與蛋白水平。 |
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370

文庫(kù)結(jié)構(gòu)
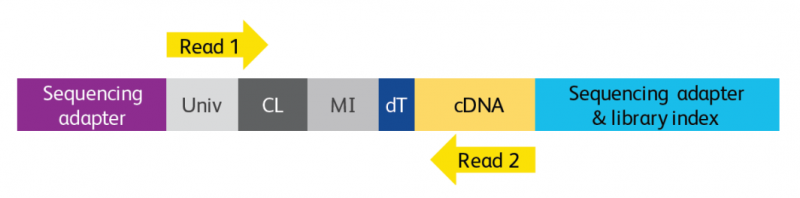
圖 BD Rhapsody 基因表達(dá)文庫(kù)示意圖
建議測(cè)序深度及參數(shù)
指標(biāo) | 參數(shù) |
測(cè)序深度 | 50,000~100,000 reads/cell |
測(cè)序類(lèi)型 | PE150 |

1、類(lèi)型:新鮮組織,原代細(xì)胞,細(xì)胞系等。
2、來(lái)源:血液提取、磁珠富集、流式富集、組織解離等。
3、樣本量及其它質(zhì)控要求:
| 樣本類(lèi)型 | 樣本質(zhì)控要求 |
| 細(xì)胞懸液 | > 10* 目標(biāo)細(xì)胞個(gè)數(shù)(底限 10,000 個(gè)細(xì)胞); |
| 活率 >80%; | |
| 濃度 500-1,000 個(gè)細(xì)胞 /ul; | |
| 細(xì)胞間無(wú)粘連(成團(tuán)率 <5%); | |
| 無(wú)大于 40um 的細(xì)胞碎片或其他顆粒物; | |
| 不存在逆轉(zhuǎn)錄抑制劑和非細(xì)胞的核酸分子。 | |
| 血液 | EDTA 抗凝的全血(不可肝素抗凝),>5ml。 |
| 組織 | 0.3cm×0.3cm(不超過(guò) 0.5cm×0.5cm)的新鮮組織,4~5 塊。 |
4、保存運(yùn)輸:
(1)細(xì)胞懸液:建議現(xiàn)場(chǎng)制備,如要運(yùn)輸,建議使用伯豪生物自主研發(fā)的單細(xì)胞保護(hù)液,4°C 運(yùn)輸,48 小時(shí)內(nèi)送達(dá)伯豪生物實(shí)驗(yàn)室。
(2)血液:EDTA 抗凝的全血,4°C 運(yùn)輸,2 小時(shí)內(nèi)送達(dá)伯豪生物實(shí)驗(yàn)室;或提取 PBMC 后凍存,干冰運(yùn)輸。
(3)組織:建議使用伯豪生物自主研發(fā)的單細(xì)胞組織保護(hù)液,4°C 運(yùn)輸,48 小時(shí)內(nèi)送達(dá)伯豪生物實(shí)驗(yàn)室。

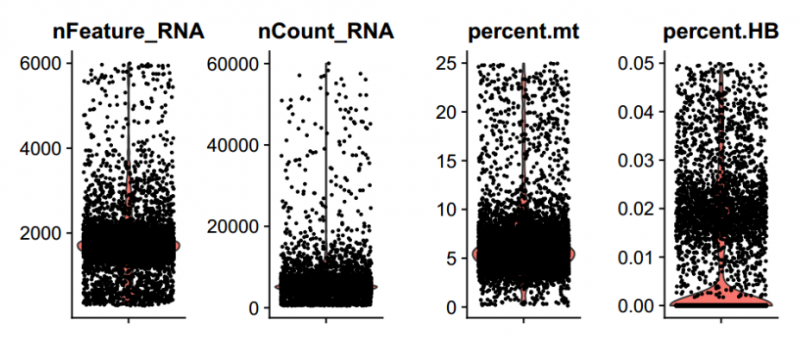
圖 數(shù)據(jù)質(zhì)控及過(guò)濾
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
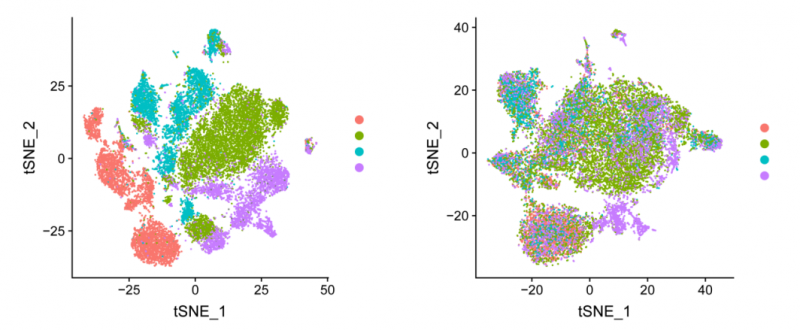
圖 去除批次效應(yīng)
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
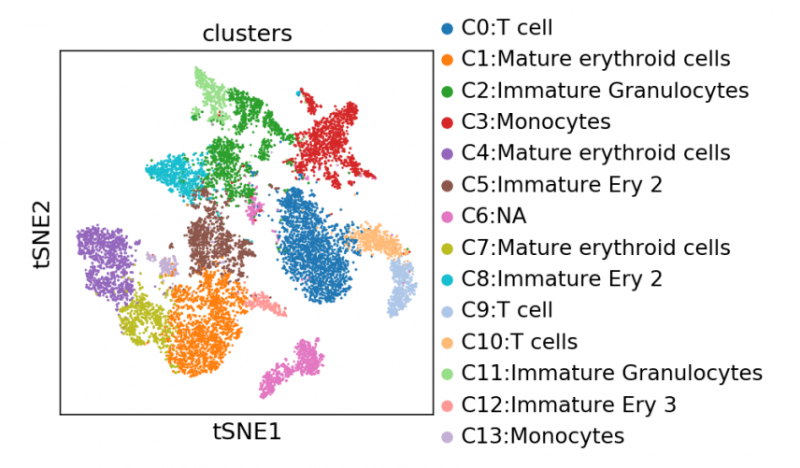
圖 單細(xì)胞 RNA 測(cè)序細(xì)胞亞群注釋
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
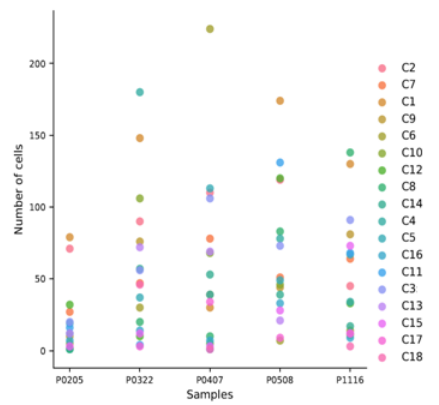
圖 單細(xì)胞 RNA 測(cè)序樣本間的細(xì)胞構(gòu)成差異
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
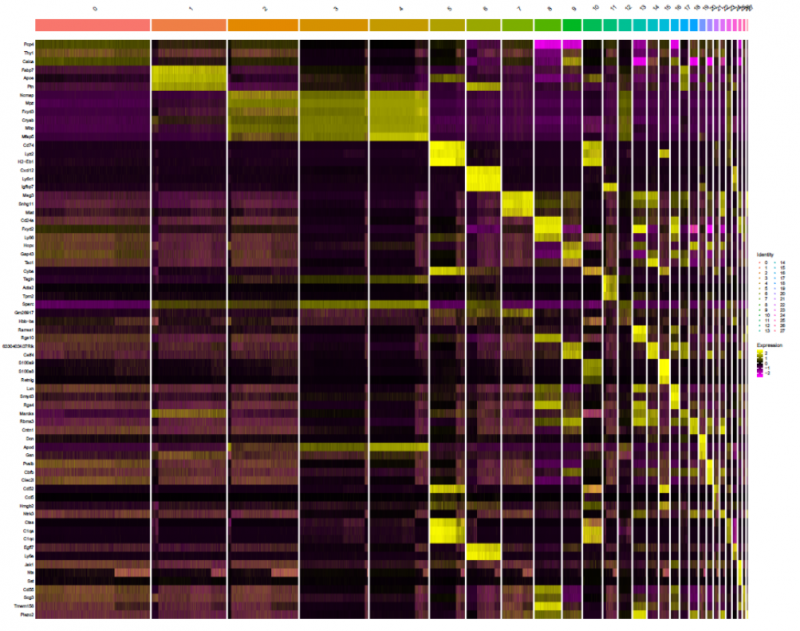
圖 單細(xì)胞 RNA 測(cè)序 Marker 基因分析
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
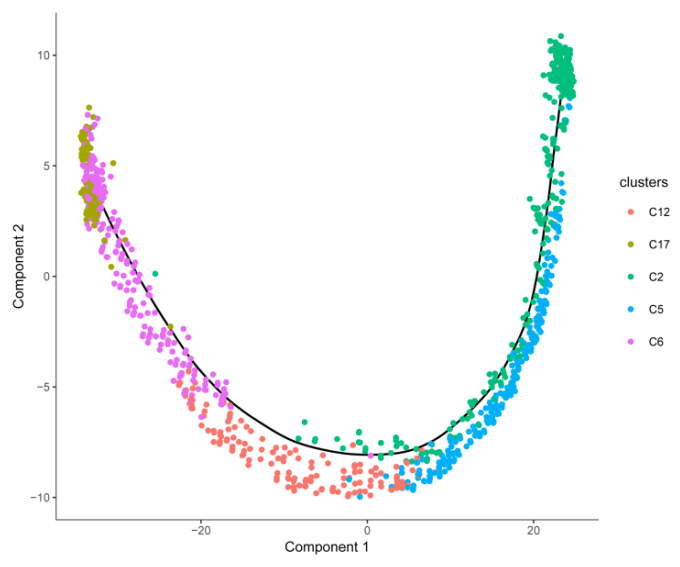
圖 單細(xì)胞 RNA 測(cè)序擬時(shí)序分析
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
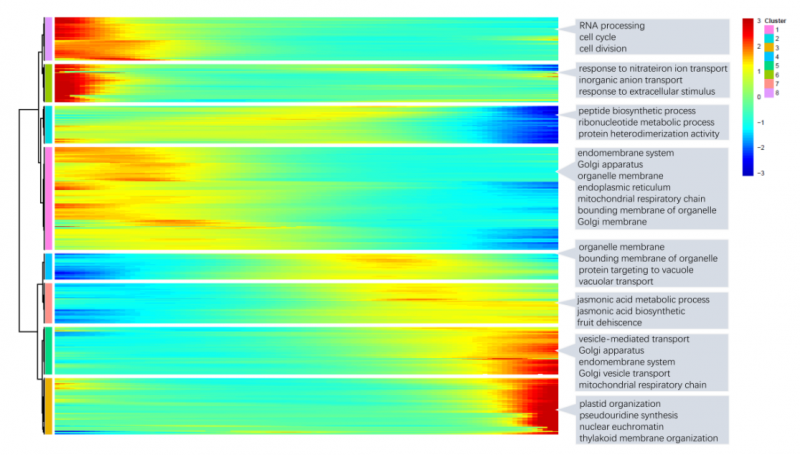
圖 單細(xì)胞 RNA 測(cè)序擬時(shí)序 GO 注釋
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
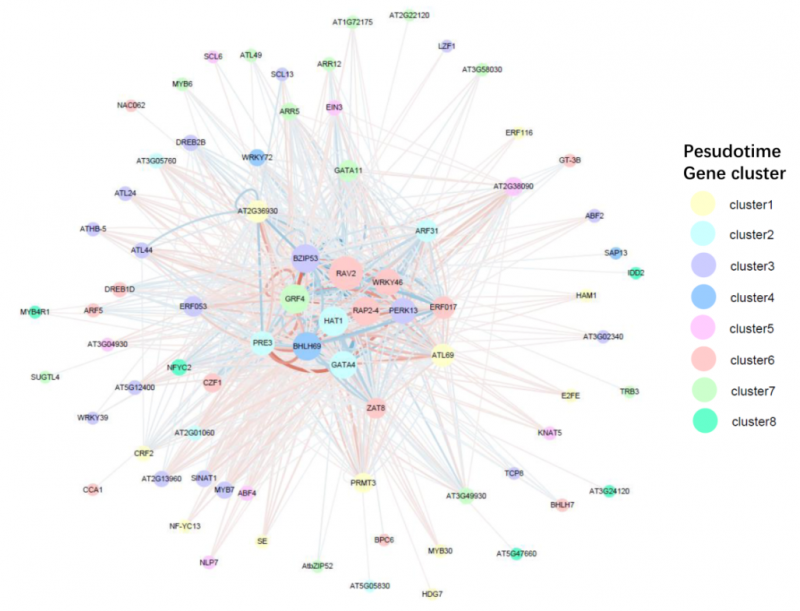
圖 單細(xì)胞 RNA 測(cè)序擬時(shí)序分化轉(zhuǎn)錄因子調(diào)控網(wǎng)絡(luò)
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
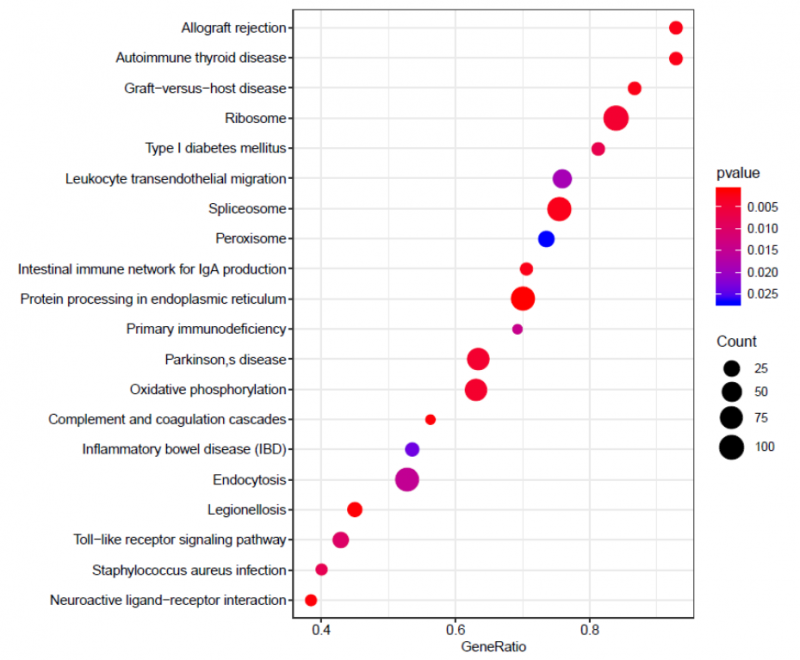
圖 單細(xì)胞 RNA 測(cè)序細(xì)胞亞群的功能富集分析
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
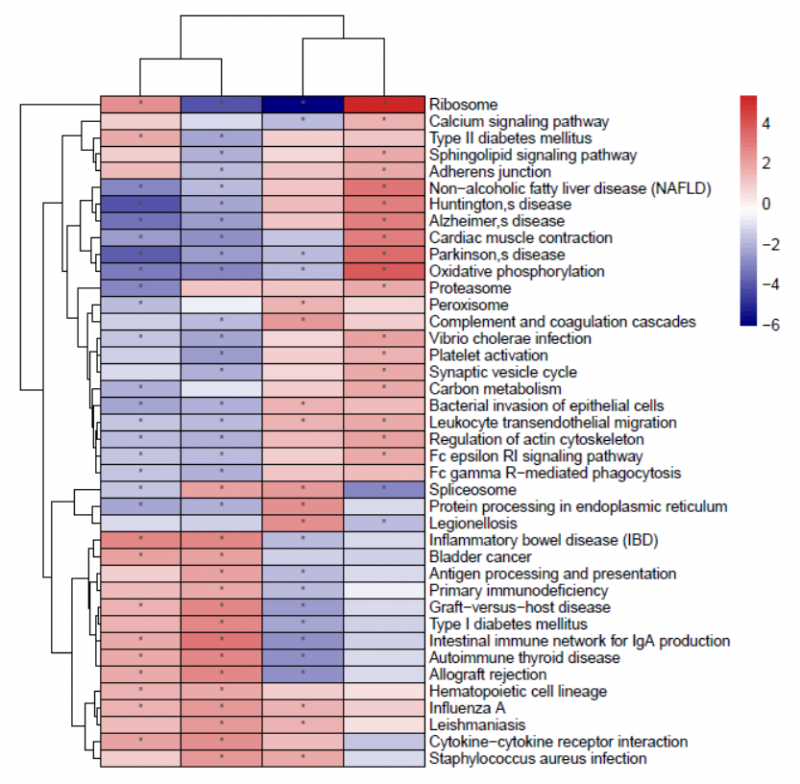
圖 單細(xì)胞 RNA 測(cè)序細(xì)胞亞群的功能富集在樣本間的差異
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
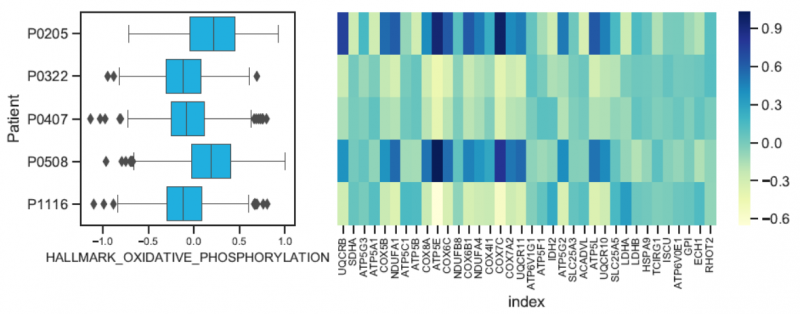
圖 單細(xì)胞 RNA 測(cè)序樣本間的功能差異分析
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
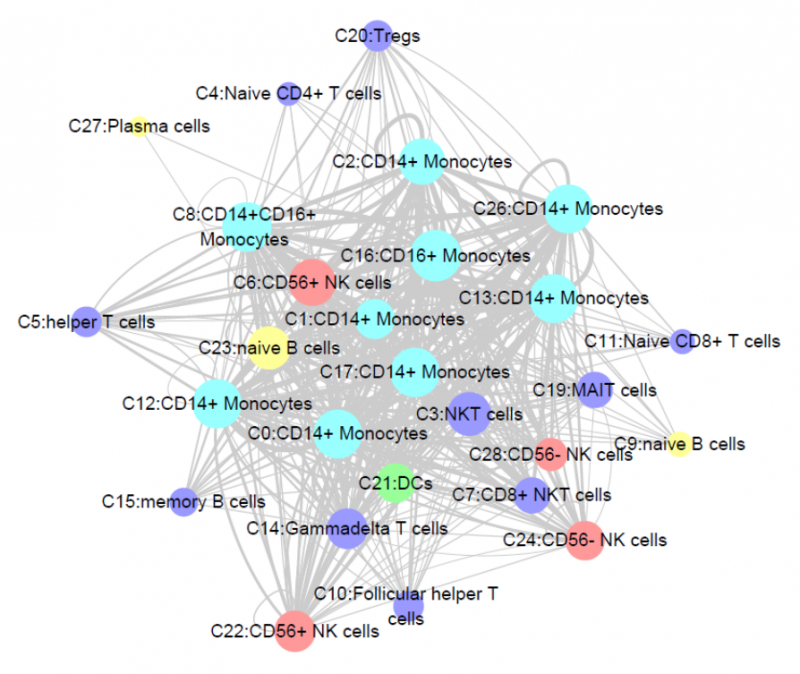
圖 細(xì)胞通訊網(wǎng)絡(luò)
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370

1、胚胎發(fā)育
人類(lèi)胚胎發(fā)育早期僅能收集微量的胚胎細(xì)胞和干細(xì)胞,這對(duì)于了解控制胚胎發(fā)育的基因調(diào)控網(wǎng)絡(luò)是一個(gè)難題,利用單細(xì)胞 RNA-Seq 技術(shù)對(duì)轉(zhuǎn)錄組進(jìn)行分析則克服了轉(zhuǎn)錄組分析對(duì)細(xì)胞數(shù)量要求的限制,近年來(lái)已有多篇相關(guān)報(bào)道。北京大學(xué)湯富酬教授課題組早在 2009 年就對(duì)單個(gè)小鼠卵裂球的進(jìn)行 RNA-Seq[4],檢測(cè)到了比微陣列技術(shù)多 75%(5270)的表達(dá)基因,并且確定了 1753 個(gè)新的可變剪接位點(diǎn)。這種單細(xì)胞 mRNA- Seq 檢測(cè)將大大提高我們對(duì)單個(gè)細(xì)胞在哺乳動(dòng)物發(fā)展中轉(zhuǎn)錄復(fù)雜性的分析能力,尤其是胚胎發(fā)育早期和干細(xì)胞這類(lèi)在體內(nèi)罕見(jiàn)的細(xì)胞群。2013 年,他們又對(duì) 90 個(gè)處于不同發(fā)育階段的人類(lèi)早期胚胎細(xì)胞以及 34 個(gè)胚胎干細(xì)胞進(jìn)行了詳盡的分析 [5],檢測(cè)出了 22,687 個(gè)母源表達(dá)基因,其中包括 8,701 條長(zhǎng)鏈非編碼 RNAs,相比于以往通過(guò) microarray 檢測(cè)出的 9,735 個(gè)母源基因數(shù)量大大增加。研究還發(fā)現(xiàn)了 2,733 條新的 lncRNAs,其中許多只在特定的發(fā)育階段表達(dá)。此外,實(shí)驗(yàn)還檢測(cè)到 1498 個(gè)基因在上胚層(Epiblast,EPI)和體外人類(lèi)胚胎干細(xì)胞間存在差異表達(dá),證實(shí)了 EPI 和 ESCs 具有顯著不同的轉(zhuǎn)錄組,有顯示差異性表達(dá),從而解答了人類(lèi)上胚層和體外干細(xì)胞之間的基因表達(dá)是否相同這一由來(lái)已久的問(wèn)題。
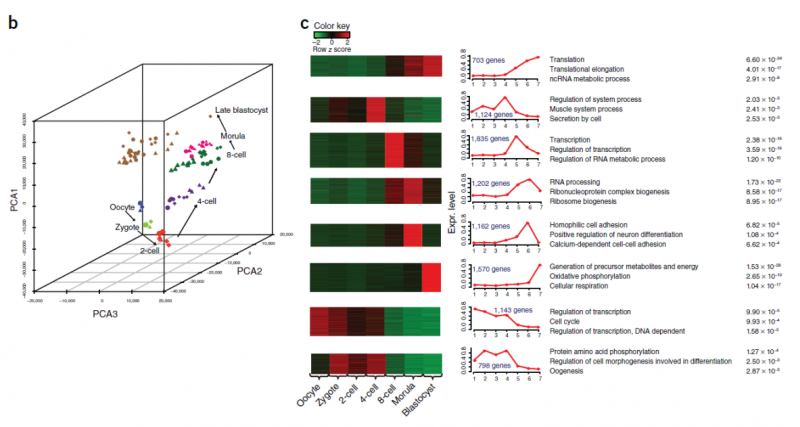
圖 單細(xì)胞 RNA 測(cè)序應(yīng)用:早期胚胎的單細(xì)胞測(cè)序 [5]
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
2、器官發(fā)育
發(fā)育是一個(gè)極其復(fù)雜的過(guò)程,器官發(fā)生涉及到多種細(xì)胞類(lèi)型的協(xié)調(diào)作用,期間基因表達(dá)受到嚴(yán)密、精準(zhǔn)的調(diào)控,這一過(guò)程中的基因表達(dá)調(diào)控機(jī)制仍然亟需深入研究。心臟是哺乳動(dòng)物在胚胎期先形成的功能器官之一。盡管已有對(duì)小鼠胚胎心臟發(fā)育的大量系統(tǒng)研究,但是人類(lèi)與小鼠的心臟存在著很大差異。2019 年,湯富酬課題組和喬杰課題組合作,利用高精度單細(xì)胞轉(zhuǎn)錄組測(cè)序技術(shù)對(duì)人類(lèi)胚胎 5 周至 25 周心臟的 4000 多個(gè)單細(xì)胞進(jìn)行了系統(tǒng)的分析 [6]。該研究系統(tǒng)鑒定了人類(lèi)胚胎期心臟的主要細(xì)胞類(lèi)型,包括心肌細(xì)胞、成纖維細(xì)胞、內(nèi)皮細(xì)胞、瓣膜細(xì)胞、心外膜細(xì)胞、平滑肌細(xì)胞以及各種免疫細(xì)胞(包括肥大細(xì)胞、巨噬細(xì)胞、T 細(xì)胞和 B 細(xì)胞)。隨著心臟發(fā)育進(jìn)展,心房、心室中的心肌細(xì)胞比例顯著下降,成纖維細(xì)胞、巨噬細(xì)胞等非心肌細(xì)胞的比例逐漸上升,這說(shuō)明在發(fā)育過(guò)程中成纖維細(xì)胞等非心肌類(lèi)型細(xì)胞對(duì)心臟發(fā)育的作用可能越來(lái)越重要。此外,還發(fā)現(xiàn)心肌細(xì)胞解剖位置特異性的基因表達(dá)特點(diǎn),而這些特點(diǎn)在早至胚胎發(fā)育第 5 周時(shí)就已經(jīng)有明確顯現(xiàn)。此外,通過(guò)與已經(jīng)發(fā)表的小鼠心臟發(fā)育的單細(xì)胞轉(zhuǎn)錄組數(shù)據(jù)進(jìn)行系統(tǒng)比較,鑒定出一系列人類(lèi)心臟發(fā)育過(guò)程中主要細(xì)胞類(lèi)型特異表達(dá)的重要基因。該研究還發(fā)現(xiàn)人類(lèi)與小鼠的心臟中心肌細(xì)胞是較為相似的細(xì)胞類(lèi)型,而成纖維細(xì)胞等細(xì)胞類(lèi)型的物種間差異較大,這為利用小鼠模型研究人類(lèi)的心臟發(fā)育提供了參比標(biāo)準(zhǔn)。
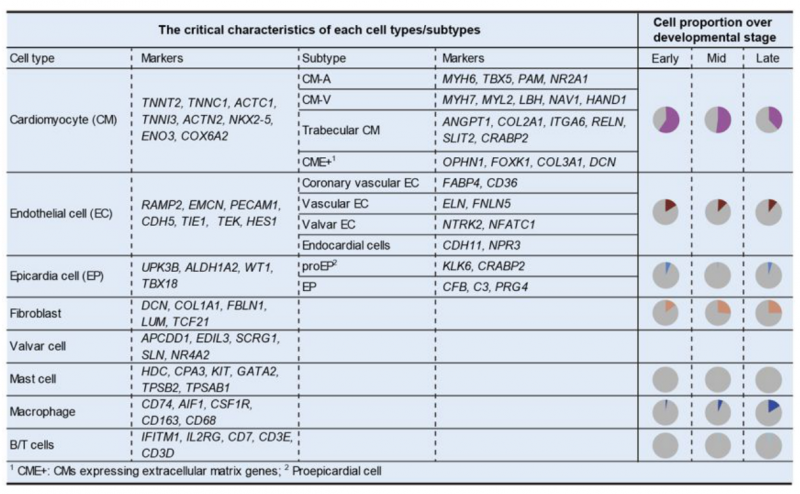
圖 單細(xì)胞 RNA 測(cè)序應(yīng)用:臟發(fā)育過(guò)程中主要細(xì)胞類(lèi)型的時(shí)空特征 [6]
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
3、干細(xì)胞分化
造血干細(xì)胞(HSC)具有長(zhǎng)期自我更新、分化成多種成熟血細(xì)胞的潛能。它起源于胚胎前體,包括生血內(nèi)皮細(xì)胞和造血干細(xì)胞前體(pre-HSCs)。2016 年,軍事醫(yī)學(xué)科學(xué)院劉兵課題組利用 pre-HSCs 特異性的表面標(biāo)志,分離出高純度的 pre-HSCs。并針對(duì) HSC 發(fā)育過(guò)程中具有代表性的 5 類(lèi)細(xì)胞進(jìn)行單細(xì)胞轉(zhuǎn)錄組測(cè)序,繪制了 HSC 發(fā)育過(guò)程的轉(zhuǎn)錄組圖譜,揭示了 pre-HSC 在轉(zhuǎn)錄活性、基因表達(dá)、代謝狀態(tài)、信號(hào)通路和轉(zhuǎn)錄因子網(wǎng)絡(luò)等方面的特征 [7]。發(fā)現(xiàn)并揭示 mTOR 信號(hào)通路在特異性調(diào)控 HSC 發(fā)育中發(fā)揮了關(guān)鍵作用。此外,還發(fā)掘出 Pre-HSC 的 98 個(gè)特征基因。該研究為今后闡明 HSC 的體內(nèi)發(fā)育規(guī)律、發(fā)掘 HSC 的體外再生策略提供了可靠的理論依據(jù)和數(shù)據(jù)資源。基于以上單細(xì)胞轉(zhuǎn)錄組數(shù)據(jù),課題組在 2019 年進(jìn)一步描繪出 HSC 發(fā)育全程的 lncRNA 動(dòng)態(tài)表達(dá)圖譜,并鑒定到在 HSC 發(fā)育過(guò)程中重要的功能性 lncRNA 分子 [8]。本研究通過(guò)生物信息學(xué)分析篩選獲得一組潛在的功能性 lncRNA,并通過(guò)體外功能實(shí)驗(yàn)發(fā)現(xiàn) 6 個(gè)可能在胚胎造血發(fā)育中發(fā)揮作用的 lncRNA。利用條件基因敲除研究策略,著重闡明了 lncRNA-H19 對(duì)于 AGM 區(qū) HSC 發(fā)生的重要作用。LncRNA-H19 的缺失使得重要造血轉(zhuǎn)錄因子(包括 Runx1 及 Spi1 等)的啟動(dòng)子區(qū)域高甲基化并下調(diào)其表達(dá),以致血管內(nèi)皮細(xì)胞向 pre-HSC 的轉(zhuǎn)化阻滯。
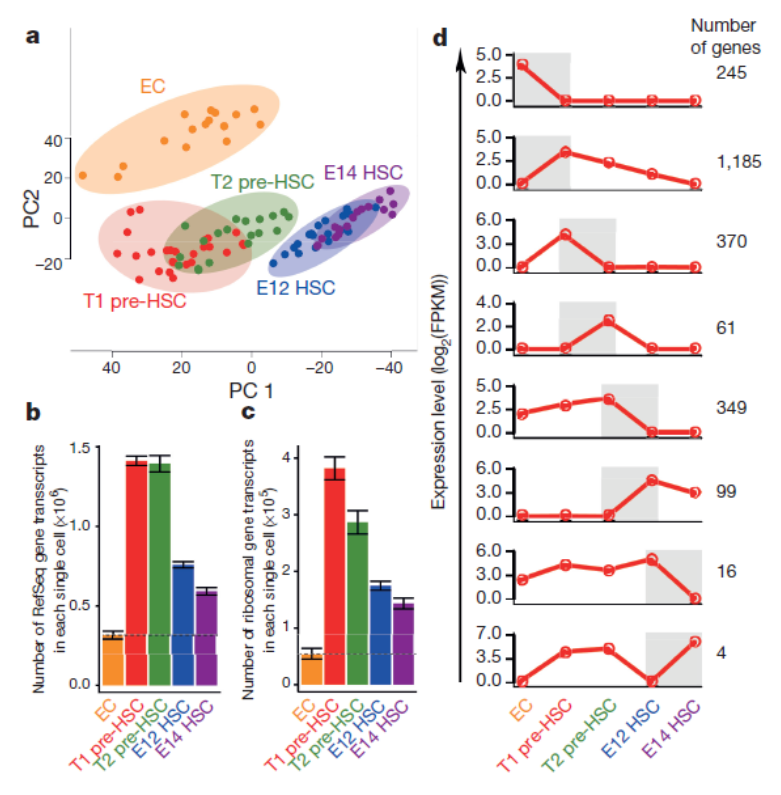
圖 單細(xì)胞 RNA 測(cè)序應(yīng)用:HSC 發(fā)育過(guò)程中的基因動(dòng)態(tài)圖譜 [7]
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
4、神經(jīng)科學(xué)
神經(jīng)科學(xué)是單細(xì)胞測(cè)序的另一個(gè)應(yīng)用領(lǐng)域,哺乳動(dòng)物的大腦中有數(shù)十億個(gè)神經(jīng)元,每個(gè)神經(jīng)元可根據(jù)形態(tài)特征,電生理特性,分子標(biāo)記進(jìn)行歸類(lèi)。在一群細(xì)胞中,單個(gè)神經(jīng)元特異性的信息就被稀釋了,少數(shù)細(xì)胞特異的基因表達(dá)則有可能無(wú)法檢測(cè)到。研究不同神經(jīng)元的表達(dá)模式能夠?yàn)槲覀兲峁└幕虮磉_(dá)圖譜、甚至表達(dá)調(diào)控網(wǎng)絡(luò)。而且,將單個(gè)神經(jīng)元的基因表達(dá)于神經(jīng)元的表型信息結(jié)合起來(lái),還能幫助我們對(duì)神經(jīng)元進(jìn)行更加準(zhǔn)確和細(xì)致的分類(lèi)。上海伯豪助力中國(guó)科學(xué)院上海生命科學(xué)研究院神經(jīng)科學(xué)研究所張旭院士研究組,通過(guò)高覆蓋度的單細(xì)胞測(cè)序和以神經(jīng)元大小為參考的層次聚類(lèi),對(duì)小鼠背根神經(jīng)節(jié)初級(jí)感覺(jué)神經(jīng)元進(jìn)行了分類(lèi),又通過(guò)全細(xì)胞膜片鉗在體記錄結(jié)合單細(xì)胞 PCR 方法可檢測(cè)各類(lèi)初級(jí)感覺(jué)神經(jīng)元對(duì)外周皮膚刺激的反應(yīng)。該工作通過(guò)高覆蓋的單細(xì)胞測(cè)序?qū)Τ跫?jí)感覺(jué)神經(jīng)元進(jìn)行了重新分類(lèi),并且建立了基因表達(dá)與在體功能的相互關(guān)系 [9]。
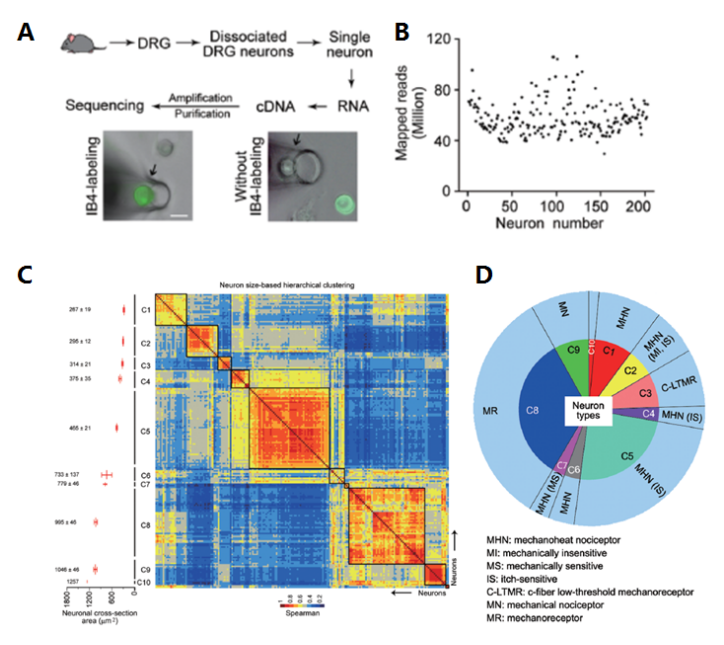
圖 單細(xì)胞 RNA 測(cè)序應(yīng)用:神經(jīng)元單細(xì)胞測(cè)序及亞群分類(lèi) [9]
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
5、腫瘤微環(huán)境
腫瘤細(xì)胞能夠采用不同策略,使人體的免疫系統(tǒng)受到抑制,不能正常殺傷腫瘤細(xì)胞,上述過(guò)程被稱(chēng)為免疫逃逸。免疫治療通過(guò)激活人體自身的免疫系統(tǒng),恢復(fù)機(jī)體正常的抗腫瘤免疫反應(yīng),從而控制與清除腫瘤。其中抗程序性死亡蛋白 1(programmed death 1, PD-1)及其配體(PD-L1)抑制劑是目前研究較多,發(fā)展較快的一種免疫療法。在各種腫瘤中,接受 PD-1 /PD-L1 抑制劑單藥治療患者的客觀有效率也僅為 10-30%,大部分患者對(duì)免疫檢查抑制劑并不敏感。隨著研究的深入,人們逐漸了解到腫瘤微環(huán)境的復(fù)雜性和多樣性,以及它對(duì)免疫治療的重要影響。腫瘤微環(huán)境與腫瘤細(xì)胞相互作用,共同介導(dǎo)了腫瘤的免疫耐受,從而影響了免疫治療的效果。抗腫瘤免疫應(yīng)答是由眾多免疫細(xì)胞和分子參與的復(fù)雜過(guò)程,受到機(jī)體復(fù)雜而精細(xì)的調(diào)控,這其中的機(jī)制仍有待進(jìn)一步研究。2017 年 6 月,北京大學(xué)張澤民課題組及其合作在 Cell 雜志發(fā)表了肝癌 T 細(xì)胞圖譜研究 [10],研究總共獲得 5063 個(gè)單細(xì)胞的數(shù)據(jù),總共聚類(lèi)為 11 個(gè) T 細(xì)胞亞群,包括 5 個(gè) CD8+ T 細(xì)胞亞群和 6 個(gè) CD4+ T 細(xì)胞亞群。其中肝癌組織中侵潤(rùn) Tregs 細(xì)胞亞群(C8_CD4-CTLA4)和耗竭性 CD8 T 細(xì)胞亞群(C4_CD8-LAYN)顯著富集,并且在這兩類(lèi)細(xì)胞中表達(dá)的 PDCD1 和 TIGIT 等,是免疫治療的靶點(diǎn)。在腫瘤侵潤(rùn)性 Tregs 中,共鑒定出 401 個(gè)特異表達(dá)基因,在耗竭性 CD8 T 細(xì)胞中,鑒定出 82 個(gè)特異表達(dá)基因,并且發(fā)現(xiàn)了新的耗竭 maker,如 LAYN,PHLDA1 和 SNAP47 等。值得注意的是,其中 22 個(gè)耗竭 marker 同時(shí)也在腫瘤侵潤(rùn)性 Tregs 細(xì)胞中高表達(dá),例如 CTLA4 和 LAYN 等。并且利用 TCGA 的數(shù)據(jù)進(jìn)行生存分析,發(fā)現(xiàn) LAYN 的表達(dá)顯著降低生存期。本研究還在單細(xì)胞水平進(jìn)行了 TCR 測(cè)序和分析,結(jié)果發(fā)現(xiàn),在腫瘤侵潤(rùn)的 exhausted T 細(xì)胞和 Tregs 細(xì)胞中,觀察到 TCRs 發(fā)生了重復(fù)使用。在腫瘤組織中含有相同 TCR 的 T 細(xì)胞比例較高,而在外周血和正常組織中比例較低,這說(shuō)明在腫瘤中的 exhausted T 細(xì)胞和 Tregs 細(xì)胞發(fā)生了克隆擴(kuò)增。并且相比于早期病人,晚期病人中 T 細(xì)胞的克隆擴(kuò)增更加明 顯。本研究為腫瘤免疫的圖譜勾畫(huà)做出了范式,也為今后其他腫瘤開(kāi)展類(lèi)似的研究及各類(lèi)腫瘤免疫的發(fā)展提供基礎(chǔ)。
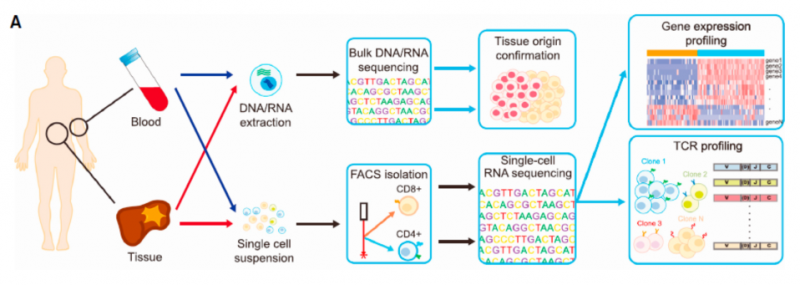
圖 單細(xì)胞 RNA 測(cè)序應(yīng)用:肝癌患者的單細(xì)胞測(cè)序 [10]
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
6、用藥指導(dǎo)
藥物誘發(fā)的超敏綜合癥(DiHS/DRESS)是一種致命的多器官炎癥性疾病,與皰疹病毒再激活和誘發(fā)的自身免疫性疾病相關(guān)。病理生理學(xué)仍然不清楚,臨床治療的選擇有限。2020 年 1 月發(fā)表在 Nature Medicine 上研究對(duì) DiHS/DRESS 患者的皮膚和血液進(jìn)行了 scRNA-seq 檢測(cè) [11]。皮膚活檢組織通過(guò)單細(xì)胞 RNAseq 發(fā)現(xiàn),患者對(duì)比健康人差異表達(dá)基因數(shù)量較多的是淋巴細(xì)胞群體。分析皮膚活檢的淋巴細(xì)胞,發(fā)現(xiàn)患者淋巴細(xì)胞呈現(xiàn)不同的分群。患者淋巴細(xì)胞中高表達(dá)的代表性基因有 CCR10,JAK3,STAT1 等。
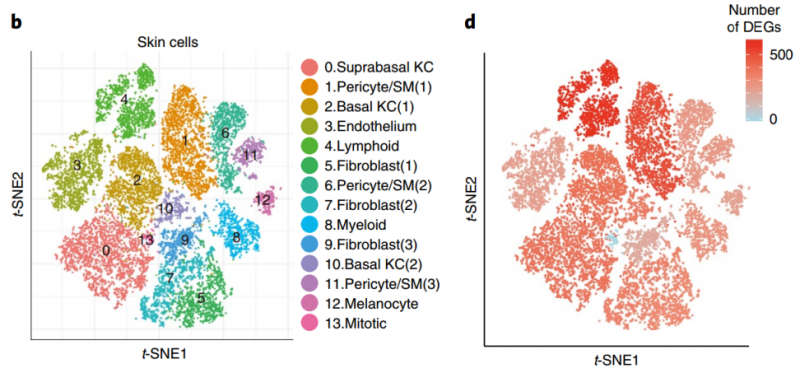
圖 單細(xì)胞 RNA 測(cè)序應(yīng)用:患者皮膚活檢的淋巴細(xì)胞呈現(xiàn)不同的分群 [11]
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
血液 PBMCs 細(xì)胞通過(guò)單細(xì)胞測(cè)序發(fā)現(xiàn),患者對(duì)比健康人差異表達(dá)基因數(shù)量較多的是 CD4 和 CD8 陽(yáng)性的亞型細(xì)胞和增殖的細(xì)胞。比較血液 PBMCs 在患者和健康人中的細(xì)胞群體分布,發(fā)現(xiàn) CCR4 和 CCR10(免疫細(xì)胞皮膚歸巢趨化因子受體)高表達(dá)的 CD4 和 CD8 陽(yáng)性細(xì)胞在患者中顯著高于健康人。在患者的這個(gè)細(xì)胞群體中,表達(dá) JAK3,STAT1,IL2RG 等基因的細(xì)胞比例明 顯高于健康人。
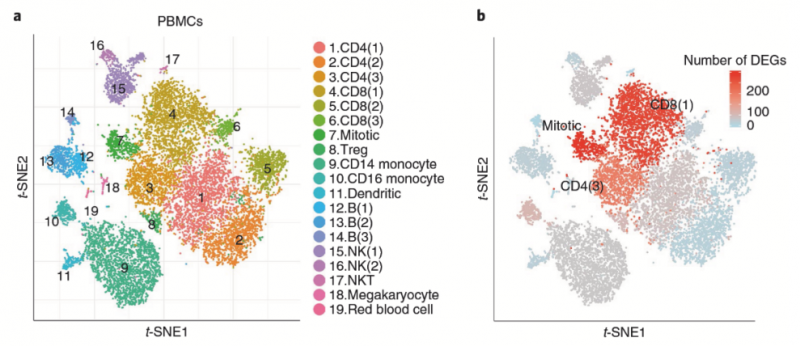
圖 單細(xì)胞 RNA 測(cè)序應(yīng)用:患者血液的 CD4 和 CD8 細(xì)胞呈現(xiàn)不同的分群 [11]
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
給予患者 JAK 抑制劑(tofacitinib)和口服抗皰疹病毒抑制劑(valganciclovir)后,患者的癥狀得到了極大的臨床獲益。治療前后的血液 PBMCs 細(xì)胞再次經(jīng)過(guò)單細(xì)胞 RNAseq 檢測(cè)發(fā)現(xiàn)細(xì)胞群體分布明 顯分化。體外實(shí)驗(yàn)也證實(shí) JAK 抑制劑(tofacitinib)和抗病毒藥物(Ganciclovir 和 Artesunate)能夠有效抑制藥物(SMX-TMP)誘導(dǎo)的 T 細(xì)胞增殖。
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370
7、病毒感染
2020 年 1 月,整個(gè)中國(guó)因新型冠狀病毒 2019-nCov 得了一場(chǎng)“感冒”。疫情一開(kāi)始,武漢病毒研究所石正麗團(tuán)隊(duì)就用實(shí)驗(yàn)證實(shí)了血管緊張素轉(zhuǎn)化酶 2(Angiotensin-converting enzyme 2,ACE2)是新型冠狀病毒感染人體的受體基因。2020 年 1 月 26 日,上海同濟(jì)大學(xué)醫(yī)學(xué)院左為研究團(tuán)隊(duì)在《bioRxiv》上發(fā)表了題為“Single-cell RNA expression profiling of ACE2, the putative receptor of Wuhan 2019-nCov”的文章 [12]。
該研究利用已有的數(shù)據(jù)庫(kù),結(jié)合單細(xì)胞 RNA 測(cè)序技術(shù)中相關(guān)生信分析,對(duì) ACE2 在人肺內(nèi)單個(gè)細(xì)胞的表達(dá)情況進(jìn)行了分析,共涉及 8 個(gè)樣本,43134 個(gè)細(xì)胞。結(jié)果表明:ACE2 受體主要在一部分(1% 左右)II 型肺泡上皮細(xì)胞(AT2)中表達(dá);同時(shí)發(fā)現(xiàn)這些 AT2 細(xì)胞除了表達(dá)病毒受體,還表達(dá)與病毒復(fù)制和傳播相關(guān)的基因,說(shuō)明其很可能是冠狀病毒的靶細(xì)胞。可見(jiàn),單細(xì)胞測(cè)序技術(shù)不僅是科研的利器,同時(shí)還為破解疫情做了應(yīng)有的貢獻(xiàn)。

圖 單細(xì)胞 RNA 測(cè)序應(yīng)用:ACE2 和其它 marker 基因在細(xì)胞亞群中的表達(dá) [12]
伯豪生物提供:?jiǎn)渭?xì)胞轉(zhuǎn)錄組測(cè)序、生信分析整體科研技術(shù)服務(wù),伯豪生物單細(xì)胞轉(zhuǎn)錄組測(cè)序科研服務(wù)商,24 小時(shí)咨詢服務(wù)熱線:021-58955370

[1] Ramskold D, Luo S, Wang YC, et al. Full-length mRNA-Seq from single-cell levels of RNA and individual circulating tumor cells. Nat Biotechnol 2012, 30(8):777-782.
[2] Picelli S, Bj?rklund ?K, Faridani OR, et al. Smart-seq2 for sensitive full-length transcriptome profiling in single cells. Nat Methods 2013, 10(11):1096-8.
[3] Picelli S, Faridani OR, Bj?rklund AK, et al. Full-length RNA-seq from single cells using Smart-seq2. Nat Protoc 2014, 9(1):171-81.
[4] Tang F, Barbacioru C, Wang Y, et al. mRNA-Seq whole-transcriptome analysis of a single cell. Nat Methods 2009, 6(5):377-382.
[5] Yan L, Yang M, Guo H, et al. Single-cell RNA-Seq profiling of human preimplantation embryos and embryonic stem cells. Nat Struct Mol Biol 2013, 20(9):1131-1139.
[6] Cui Y, Zheng Y, Liu X, et al. Single-Cell Transcriptome Analysis Maps the Developmental Track of the Human Heart. Cell Rep 2019, 26(7):1934-1950.e5.
[7] Zhou F, Li X, Wang W, et al. Tracing haematopoietic stem cell formation at single-cell resolution. Nature 2016, 533(7604):487-92.
[8] Zhou J, Xu J, Zhang L, et al. Combined Single-Cell Profiling of lncRNAs and Functional Screening Reveals that H19 Is Pivotal for Embryonic Hematopoietic Stem Cell Development. Cell Stem Cell 2019, 24(2):285-298.e5.
[9] Li CL, Li KC, Wu D, et al. Somatosensory neuron types identified by high-coverage single-cell RNA-sequencing and functional heterogeneity. Cell Res 2015, 26(1):83-102.
[10] Zheng C, Zheng L, Yoo JK, et al. Landscape of Infiltrating T Cells in Liver Cancer Revealed by Single-Cell Sequencing. Cell 2017, 169(7):1342-1356.
[11] Kim D, Kobayashi T, Voisin B, Jet al. Targeted therapy guided by single-cell transcriptomic analysis in drug-induced hypersensitivity syndrome: a case report. Nat Med 2020,26(2):236-243.
[12] Yu Zhao, Zixian Zhao, Yujia Wang, et al. Single-cell RNA expression profiling of ACE2, the putative receptor of Wuhan 2019-nCov. bioRxiv 2020.

| 序號(hào) | 文件類(lèi)型 | 查閱 |
| 1 | 【畫(huà)冊(cè)】單細(xì)胞全長(zhǎng)轉(zhuǎn)錄本測(cè)序解決方案 | 點(diǎn)擊查看 |
| 2 | 【畫(huà)冊(cè)】石蠟樣本(FFPE)單細(xì)胞轉(zhuǎn)錄組測(cè)序解決方案 | 點(diǎn)擊查看 |
| 3 | 【畫(huà)冊(cè)】單細(xì)胞測(cè)序解決方案 | 點(diǎn)擊查看 |

